

A medium-sized sparrow, the American Tree Sparrow summers in the near-perpetual sunshine of northern Canada and Alaska, moving into much of the northern United States during the winter.
Breeding and wintering ranges of the American Tree Sparrow do not overlap, so all individuals are migratory. Migration takes place mostly at night, and often in flocks. Male American Tree Sparrows often winter farther north than females.
American Tree Sparrows winter throughout much of the northern and central U.S., where winters are often cold and snowy. They forage by scratching around on the ground for seeds, but when there is snow covering the ground they are also known to beat their wings against weeds and then eat the seeds that have fallen onto the snow.
Photograph © Mr. Lawton.
BREEDING MALE

Photograph © Glenn Bartley.
Length 6 in. Wingspan: 9 in.
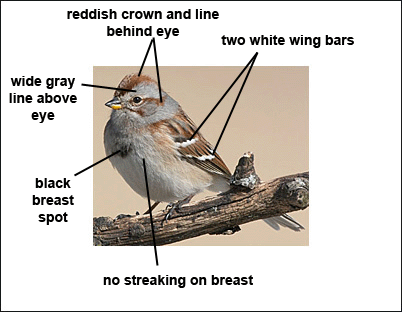
- Reddish-brown wings with two white wing bars.
- Gray head with rufous eyeline and crown.
- Dark spot in center of breast.
- Upper mandible is dark, while lower mandible is yellow.
Female
Same as male.
Seasonal change in appearance
Become generally buffier in appearance. Rufous on crown may split into a central stripe.
Juvenile
Resembles adult but has a heavily streaked breast and face.
Habitat
Nests in Arctic scrub, and winters in weedy fields and along brushy roadsides.
Diet
Primarily seeds in winter and insects in summer.
Behavior
Forages on the ground or in low bushes, often in flocks in the winter.
Range
Widespread breeder in Alaska and across Canada north of the southern provinces. Winters across much of the U.S. population appears stable, perhaps due to largely isolated breeding habitat.
More information:
Bent Life History
Visit the Bent Life History for extensive additional information on the American Tree Sparrow.
Fun Facts
Despite its name, much of the breeding range of this species is north of the treeline.
Vocalizations
Song consists of a few clear notes followed by a high warble. Call is a soft teedle-ee.
Similar Species
Field Sparrow

The Field Sparrow lacks a central breast spot, and has a pink bill.
Chipping Sparrow

Photograph © Glenn Bartley.
Chipping Sparrows lack central spot, has more distinct eyeline and white above the eye.
Nesting
An open cup of plant material, sometimes lined with ptarmigan feathers, usually placed on or near the ground beneath a shrub.
Number: Usually 5, but can range from 4 to 6.
Color: Pale blue or green with brownish markings.
Incubation and fledging:
- Young hatch at about 11 to 13 days.
- Young fledge at about 8 to 10 days, receive care from parents for another 3-4 weeks.
Bent Life History of the American Tree Sparrow
Published by the Smithsonian Institution between the 1920s and the 1950s, the Bent life history series of monographs provide an often colorful description of the birds of North America. Arthur Cleveland Bent was the lead author for the series. The Bent series is a great resource and often includes quotes from early American Ornithologists, including Audubon, Townsend, Wilson, Sutton and many others.
Bent Life History for the American Tree Sparrow – the common name and sub-species reflect the nomenclature in use at the time the description was written.
SPIZELLA ARBOREA (Wilson)
Contributed by A. MARGUERITE BAUMGARTNERHABITS
To anyone who has tramped our snowy fields and brushy hedgerows in winter, the little tree sparrow is a familiar and beloved bird. One of the most abundant winter residents over much of the northern and central United States, its gentle confiding ways and cheerful warblings have ever endeared it to those who go afield with binoculars or who feed the winter birds about their gardens. Easily identified by its pert chestnut cap, clear gray breast with a single black “brooch,” white wingbars, and a bill dark above and yellow below, the tree sparrow is most frequently confused with its relatives, the somewhat similar but smaller chipping and field sparrows.
Two geographic races are now recognized, the nominate eastern Spizella arborea arborea and the western form, S. a. ocliracea, with a wide belt of intergradation between them through the Prairie Provinces and the Plains States. With no perceptible differences in measurements or in the juvenal plumage, typical ocltracea differs from the eastern race primarily in degree of color, having a slightly paler back, a richer wash of ochraceous buff on the sides, and a heavier veiling of buff on the crown. Although the line of demarcation between the two falls in mid-continent in winter, during the breeding season the eastern race sweeps far westward. It nests from the Atlantic coast to Anderson Valley, almost to the boundary of the Yukon Territory (Preble, 1908), while ochracea breeds across northern MacKenzie and the Yukon, throughout Alaska as far north as there is any scrubby growth, and down the mountain ranges of British Columbia in the sparse forests just below timberline.
The tree sparrow is another example of a misnomer by our pioneer forefathers, for few birds spend less time in trees, either in winter or summer. The early settlers saw in it a superficial resemblance to the chestnut-capped tree sparrow (Passer morttanus) of Europe and, either in ignorance of specific differences or in nostalgic recollecton, called this little American sparrow by the same name. Actually our tree sparrow might more accurately he called brush sparrow. for its haunts include the scrubby edges of our fields and marshes, hedgerows, and fallow fields. In summer it travels far beyond the dense Canadian woodlands to the lake-dotted open forests of the upper Hudsonian Zone. Though it nests occasionally in the dense, stunted spruces characteristic of this region, it spends most of its time in the brushy willows and birches that fringe the pools and boggy meadows, and the overwhelming majority of nests on record have been on the ground.
An analysis of hundreds of faunal works, local lists, Christmas censuses, and field notes, shows the center of abundance of tree sparrows in a normal winter to be through central Nebraska, Kansas, Iowa, and Missouri, and across the corn belt to the middle Atlantic States (Baumgartner, 1939). An abnormally mild season finds a greater number wintering in the northern states and southern Canada, while early storms and excessive snows drive them farther south. In Oklahoma, near the southern border of their regular range, I have found their occurrence as variable as Oklahoma weather. In Tennessee, where they occur only rarely, they are generally associated with low temperatures and a blanket of snow (Laskey, 1934b).
Winter distribution of species: In the course of two seasons of collecting at Ithaca, N.Y., it was discovered that the proportion of males ran considerably higher than females during the nonmigratory period, which I arbitrarily limited to the dates December 20 to March 1. In fall and spring the female population equaled or exceeded that of males. Postulating that this indicated segregation of the sexes during the winter, I wrote all the leading museums in the country for lists of their tree sparrow specimens, in the hope of plotting the respective ranges. While many did not have numbers adequate for any conclusions, the larger series corroborated the figures from my personal collecting and banding (indicated in the following list by *), showing that males tend to winter farther northward and females farther southward:
Part of Range of Number of Winter Species MalesFemales Locality Northcrn Ontario 16 0 British Columbia 11 2 Idaho 16 9 Michigan 28 5 *New York (1933: 35)102 31 Connecticut 21 1 Central *Indiana (1935) 16 22 Pennsylvania 26 27 District of Columbia 17 17 Kansas 185 158 Southern *Oklahoma (1940-41)31 78 *Oklahoma(1960) 40 113
During late February and March of 1960 Oklahoma had a period of severe cold and heavy snows which brought in an unprecedented number of tree sparrows. During a single month I handled almost 200, most of which rel)cated several times a day. When my hands were not too numb nor my glasses fogged, I noted external sex characteristics (see Plumages), and recorded a tentative sex identification. While some were not differentiated sharply enough even to hazard a guess, I have 113 cards marked as unquestionably female with narrow crowns heavily streaked with dark shafts, and 40 possibly immature males or adult females, the group most difficult to sex by external criteria. Of the 168 tree sparrows I banded between 1948 and 1959, I designated 110 as females, the rest an indetermined proportion of adult females and males.
Migration: Between the winter range and the breeding grounds of the tree sparrow stretch almost 1,000 miles of territory in which, the species occurs only as a migrant. Alaskan birds must travel some 3,000 miles to reach their destination, but most of the birds that winter through central United States may be assumed to journey between 1,500 and 2,000 miles to nest.
In Oklahoma, in the southern part of their range, the first evidence of spring movement comes in late February, and the majority are normally gone by mid-March. Following the severe storms of March 1960, new birds continued to visit my banding station at Stillwater until March 26, and a dozen or more individuals remained until April 5.
In the Mississippi Valley, Cooke (1888) found the crest of the migration to pass between March 30 and April 8. Eaton (1910) lists migration in New York State chiefly through late March and the first three weeks of April; the latest is May 15 at Ithaca (Edminster, MS.). Traveling primarily by night, flocks of five or ten or hundreds arrive in waves, remain a few days or weeks, and drift away a few at a time.
The sequence of migrants and winter residents as presented by W. P. Smith (1926), Homey (1926), and Austin (1932b), agrees substantially with the movements observed at Ithaca, N.Y. Five years of records from Dr. A. C. Frazer’s station and a check group of 54 regular winter residents at my own traps showed parallel behavior: A sprinkling of the winter residents (birds banded between December 20 and March 1) disappeared during mid-February, which probably represents local wandering rather than true migration. D tiring the third week of February a number of new birds were banded and a few more winter residents drifted away, consistently enough at both stations to suggest the vanguard of true migration. March saw a steady increase in migrants and departure dates of winter residents with a pronounced crest the 4th week. Smaller crests occurred in mid-April for migrants, while the number of winter residents decreased steadily through the mouth. Interestingly, the last birds to leave on May 1 were on of my winter “standhys” and a winter resident from the Frazer station.
The proportion of the sexes during spring migration indicates that these birds do not travel as mated pairs. In late February at the end of the winter period, males constituted 90 percent of the birds collected from a flock near Ithaca. Thereafter the percentage of females increased steadily until it equaled the males in late March, the crest period for departing winter residents and incoming migrants. During the mid-April crest, proportions were nearly equal again with 45 percent male birds, many of which upon dissection showed evidence of immaturity. On April 26 the last collection netted 86 percent females and a sprinkling of small males, presumably first year birds (Ileydweiller, 1936).
Nesting: At the beginning of my graduate study on the life history of the tree sparrow, I wrote hopefully to Mr. Bent for any available information on the nesting habits of this species. He replied that there was little beyond locality records and descriptions of nests and eggs, adding tersely, “It is regrettable that you did not select a bird you could study in the field yourself.” The following account is the outcome of that brief comment (Baumgartner, 1937b):
In the wet, brushy wastes of the Canadian Northland, beyond arable land or usable timber, from the northern third of the Hudsonian spruce -timber as far north as there is any scrubby growth, the tree sparrows find optimum conditions for nesting and for rearing their young. Churchill, Manitoba, at latitude 580 N. hallway up the west coast of Hudson Bay and 5 miles beyond timberline, proved an ideal locality to study the nesting habits of this species. Connected by rail with the outside world in 1929, it was then the only spot that could be reached early enough in the season to trace the complete nesting cycle without spending a winter in the North. The embryo town was hospitable, and the nearby river flat, a bouldery, hummocky, pool-dotted stretch of tundra extending back to timberline, was interspersed with patches of scrub birch and willow, a maximum of 4 or 5 feet high, in which tree sparrows were one of the most abundant sl)eeies.
During 5 months spent on the nesting grounds: throughout June and July in 1933 and from June 4 to August 21 in 1934: I found a total of 26 nests, 9 of which ~ ere kept under constant observation from the time of their discovery. While the birds were common, their nests, usually hidden in the densest tangles of the scrubby thickets, were difficult to find. rrhe nests studied, however, showed such uniformity that I was satisfied they were both normal and typical for the species.
In 1946 Walkinshaw (1948) gathered nesting data on the western tree sparrow in western Alaska. His observations and data are remarkably consistent with my Churchill records and corroborate the close relationship of the two forms.
According to Taverner and Sutton (1934) the tree sparrow spends about four months at Churchill, arriving about May 25, and is seen last on September 29. The occasional few of the first two days (Sutton’s field notes) were followed almost immediately by enormous numbers of both sexes, scattered (none in flocks) through the stunted willows of the river flats. In Alaska, Nelson (1887) likewise noted that flocks were broken up when they reached the breeding grounds and that the birds were mated soon after.
When I arrived at Churchill in early June, the tree sparrows were already established on their territories and had begun nest building. The successive stages of the reproductive cycle and their inclusive dates at three typical nests were:
Dates of Period Activity *
Nest I Nest III Average Length Nest IX of Period Ncst building
Resting Laying
Incubating
Hatching
Young leaving nest
Parents feeding fledglings Molting
Fall flocking (*At Churchill, Completed June 5 5 days June 11 to 16 (5 eggs)
Completed June 13 2 days June 16 to 21 (6 eggs)
June 16 to 28 June 20 to
July 3 1 at 4 p.m., 3 by 10 am., June 28; 2 by 2 p.m.
3 by 7 a.m. July 3 June 29 last left at noon July 8 3 on July 11 (disturbed), 2 on July 13 7 days (at nest II)
2 to 3 days June 25 to 28 4 to 6 days, (4 eggs) usually 1 egg each day 12 to 13 days all within 7 to 30 hours June 27 to July 10 1 at 9 am., 1 at 8 pin.
July 9,1 by noon July 10 only 1 left, on July 18 9 to 0~ days (8 days if disturbed)
2 weeks Until first week of August (a few to August 19)
Throughout August by both adults 3 weeks and young First seen August 17 and 19 1 month Manitoba; nest dates in 1933, August data in 1934.)
In the short northern summer it is highly unlikely that more than one brood is reared each year. As the earliest young at Churchill left the nest on July 8 and both parents continued to feed them for another two weeks or more, a second brood could not have been started before July 22. As it takes some 70 days after egg-laying to bring young through the postjuvenal molt., such a late brood could hardly be ready to migrate by September 29, the latest record for t.he species’ occurrence there.
Song is the tree sparrow’s chief asset in courtship display, for the species is plain colored and the sexes are alike. In courting either sex may be the aggressor, pursuing the other and flaunting its humble beauties. Of one pair observed for some time and finally collected, the pursued bird proved to be the male. One nest-building female approaching her nest with a ptarmigan feather was filled with a sudden excitement when her mate came to sing nearby. She dropped the feather, hopped to a twig about a foot from the ground, spread and fluttered her wings, and uttered an alluring “wehy-weliy-wehywehy.” The male sang on indifferently, and she picked up her feather and went onto the nest.
In another case the female fed quietly on the ground while her suitor, sitting on the bush above her, uttered a rapid series of stacatto chips, puffed his plumage, spread his wings, and then darted to the ground with much fluttering. In acknowledgment she preened daintily, fluttered her wings with neck much extended, and murmured the soft “wehy” notes which brought him to her. Fluttering over her for an instant, he copulated and darted back to his perch. He repeated the procedure several times in the next two minutes. There was no singing at this time. Sometimes copulation took place without display of any kind, the male simply flying to his mate while she was feeding or at the nest. The nest I pair were seen to copulate at the nest on three occasions, once during its building, twice within half an hour the clay before the laying of the second egg.
The extent of the tree sparrow’s summer territory proved to be considerably smaller than the winter feeding range, as anticipated. Two nesting areas were studied in detail: a stretch of open tundra bordered on one side by a small patch of brush, near which was a single nest (nest I): and a tangled thicket of low willows and birches, extending some 300 by 800 feet along the river fiats, within whose confines dwelt four families in 1933 (nests Ill: VI) and three in 1934 (nests C, D, E).
Though I marked no adults the first season for fear of upsetting their natural behavior patterns, I could tell the males apart by their song patterns, and the females by watching them as they left the nest. In 1934 I decorated a few adults with gay chicken feathers to corroborate the first year’s observations. Young birds were banded, but the tailless, soft jur-enal plumage would not hold additional feathers (Heydweiller, 1935).
The size of individual territories was found to vary with the season and the location. During the first week in June, when birds were engaged chiefly in feeding together, courting, and preliminary building, territories were large and poorly defined. One male, feeding with his mate at a little pooi, made vicious attacks upon a third bird and pursued it 400 to 500 feet before circling back. Then abruptly the pair flew ~ mile downstream and disappeared in the brush.
With the onset of incubation, areas became more limited. Isolated nest I was surrounded by a territory some 600 by 700 feet in diameter, and both birds frequently fed and later obtained food for their young at a drainage ditch 450 feet from the nest. In t.he more congested thicket occupied by nests Ill: VI, the song perches of the various males encompassed territories only 200 to 300 feet in diameter. Occasional overlaps occurred without friction, provided the other male was singing at the opposite side of his territory; and in spite of close quarters very little fighting was observed.
While the young were in the nest, both parents foraged in the thick tangle of low leafy branches of the tiny birches and willows, frequently only 10 or 15 feet from their charges. Throughout the latter part of July 1934, the gay plumes with which the adults were marked waved from the same tiny territories while the fledglings scuttled among the underbrush.
During early August., when the young were fully fledged and able to feed themselves, the three 1934 broods in this area ranged freely throughout the 300- by 500-foot patch of thicket, though they still maintained family unity. Not until the 19th of August did they emerge from the thick cover and merge with the flocks that were gathering in the grasses and moss of the river flats.
The main defense of territory against invasion is song, strengthened by an occasional brisk assault upon an intruder. While this is usually by the male, his mate occasionally assumes the offensive. At nest I when the young were a day old, the brooding female suddenly darted from the nest and made a vicious attack upon another tree sparrow. At nest IV the incubating female flew at a yellow warbler that perched carelessly some 20 feet from her: her mate was singing only 150 feet from the nest, but she apparently considered the case too urgent t.o wait for him to expel the trespasser.
At my intrusion the parent birds chipped anxiously, but neither attacked, as will hawks, terns, owls, and jays, nor feigned injury as did their neighbors the shore birds, though if followed they would guilefully lead me farther and farther afield. The chief protection of the incubating female was her streaky coloration, which she relied on until almost stepped upon.
Specific sites for the 26 nests I studied at Churchill were of three types:
1. Dwarf willow: birch thickets: 20 nests in tussocks of grass or depressions in the ground at the base of shaggy bushes, concealed by dry grasses and dead branches.
2. Open tundra, near patch of brush: four nests in depressions in mossy hummocks, concealed by grasses or semiprostrate shrubs.
3. Spruces at timberline: one nest at base of stunted spruce on open tundra; one nest 5 feet up in a small spruce under overhanging branches, built on the remains of an old redpoll nest.
All but the last of these were on or within a few inches of the ground. MacFarlane (1891) who examined more than 200 nests in the Anderson River valley, found most of them on the ground, but a few in dwarf willows 1 to 4 feet high.
The nest is usually built in three layers: an outer shell 20 to 40 millimeters thick, composed of heavy grass and weed stems, rootlets, bits of moss, lichens, shreds of bark, and an occasional leaf or twig; an inner coat of fine, dry grass stems, about 10 millimeters thick; and finally a soft lining of ptarmigan feathers. Generally the nests are compact, well-built, rather heavy, and open to the sky, though a few open at the side. One found in a deeper depression lacked the outer shell almost entirely and was lightly and loosely woven. The lining feathers, ranging from 29 to 152 in number, were frequently woven into the inner shell. Other materials than ptarmigan feathers may be used for linings: five nests had pintail feathers, one of them exclusively. Another had dog hairs interspersed among the feathers. One nest contained a soft mass of the thin, wiry mosses that carpet the ground. The presence of a small patch of lemming fur in the bottom of another suggests the birds sometimes raid this animal’s old winter nests that are found everywhere above the ground. One enterprising bird that built near the railway station used shreds of waste cloth.
The average dimensions of tree sparrow nests at Churchill were as follows:
ExteriorInterior diameterdiameter Interior Depth (top, in(top, in depth (in overall (in Weight millimeters)millimeters) millimeters) millimeters) (in grams)
Average 130 50 35 54 15. 6 Maximum 150 55 45 70 25. 9 Minimum 110 40 30 45 4. 9
Nest building is by the female alone, but her mate takes an active interest in her activities and is never far away. Singing a few feet above her while she works at the nest, he frisks dutifully after her when she goes for another feather or straw, then back to his perch when she returns, or sometimes down to the nest to mate with her. The source of the material is never far distant, for grasses and moss are the stuff of the tundra itself, and ptarmigan feathers are scattered everywhere as these birds shed their winter dress. Building is a leisurely process, and after three or four trips for material, the pair usually ifits off to feed about the marshy edges of a pooi, and the observer might sit and shiver for 2 hours or the rest of the day before they return.
The construction of a nest observed almost from its beginning was briefly as follows:
June 5 and 6: Pair seen feeding together; no indications of building though territory defined, as indicated by feeding range and attacks on others of this species.
June 11, 8:30 a.m.: Nest found, barely started, a small depression in the damp ground with moss padded partly across the back, bits of straw thinly woven across the front, and two or three soggy bits of dead grass on the floor. Female came to nest three times in 2 hours. 3:00 p.m.: Floor sparsely laid with soggy straws from the front to one-third of way back.
June 12, 7:30 a.m.: Nest much more rounded and built up in front. Yesterday’s flooring covered with bits of moss and peaty sod. 7:30 p.m.: Rim built up an inch above yesterday’s mark and broader, now 3~ inch thick in back as well as front of nest. Floor well padded with moss and sod. Length of time for building outer shell approximately 3 days.
June 13, 3:00 p.m.: Nest has a beautifully woven inner coat of fine dead grass, stems, and one ptarmigan feather. Inner coat practically completed in % day.
June 14, 6:15 p.m.: Three feathers in nest.
June 15, 7 p.m.: Ten more feathers and finer grasses added to inner shell, burying one feather.
June 16, 10:30 a.m.: One new feather.
June 17, 18, and 19: No change, nest looks complete. Lining completed in 3 days, followed by a rest of 5 days. When I examined this nest at the end of the season there were 43 feathers in it.
June 21, 12:30 p.m.: One egg.
Eggs: The tree sparrow usually lays from three to five eggs, occasionally six. They are ovate with some tending toward short ovate, and they are slightly glossy. The ground is pale bluish or pale greenish-white, profusely spotted and speckled with “wood brown,” “cinnamon brown,” or “auburn.” These markings are generously scattered over the entire surface, often entirely obscuring the greenishwhite ground and giving it a light brown appearance. In addition to the heavy speckling there are frequently a few long thin black scrawls.
The measurements of SO eggs of S. a. arborea average 19.5 by 14.3 millimeters; the eggs showing the four extremes measure 20.8 by 15.0, 17.7 by 14.5, and 18.3 by 13.5 millimeters. The measurements of 40 eggs of S. a. ochracea average 18.8 by 14.0 millimeters; the eggs showing the four extremes measure 21.3, by 15.2, 16.8 by 13.7, and 17.3 by 13.2 millimeters.
In color, size, and shape, the tree sparrow eggs at Churchill displayed a striking variation, not only in the different nests, but within a single set. Ground color ranged from pale blue to greenish, and from a clear, almost bright hue to a dull, leaden appearance. In some, the brown fieckings were so evenly washed over the whole surface that the ground color was almost obliterated, though generally they were more concentrated at the large end of the egg. The shape has been described as oval, ovate, or ovoidal. Frequently one egg in a nest was conspicuously more rounded than the others.
The following measurements were made during early incubation:
Length Width (in milli-(in miii- Circumference Weight meter.s) meters) (in millimeters) (in grams) Average 19.1 14.2 44.0 2.12 Maximum 21.0 15.5 4& 0 2.40 Minimum 17.5 14.0 41.0 1.82 Variation in one eeL 21. 0 14. 5 long and oval 2. 40 (Nest VI) 19.5 14. 0 normal shape 2.08
The average egg weight equals 11.37 percent of the yearly average weight of the female bird (18.62 grams). At nest I weights were taken at intervals of 3 or 4 days to check the percentage of loss. Total loss was 0.30 gram, or 14.63 percent.
A normal clutch of the tree sparrow at Churchill ranged from four to six eggs; the majority consisting of five. MacFarlane’s (1891) records for 216 nests were four to five, with occasional sets of six or seven. Sets of two and three are apt to be incomplete. In order to determine the capacity for laying of this species, the third egg at nest II was removed each morning. Unlike the famous flicker that laid 71 eggs, the female settled down to incubate after her fifth egg and laid no more. The experiment was not repeated, and may represent only an individual case.
Of 66 eggs in 14 nests that were watched through an undisturbed hatching, five eggs or 7 percent did not hatch and presumably were infertile. Of these, three belonged to a 6-egg set, and one to a belated 4-egg set.
After the nest was completed, the birds usually rested for several days, during which they seldom visited the nest. Thereafter an egg was laid daily. At five nests observed, only one bird skipped a day (between the second and third eggs). The eggs were laid in the early morning, between 6:00 and 7:30 am. At nest I the second egg had not been deposited at 6:45, but was laid before I returned at 7:45. On the 4th day the new egg was in the nest when I arrived at 6:30.
During egg laying the birds show somewhat more interest in the nest. In 10 visits to nest I during this time, I found the female covering the eggs on two occasions, once from 5:30 to 6:00 a. in., returning at 6:25 to lay, and once in mid-afternoon. In addition the male visited her twice, and came once when she was absent. Other females were found on their nests at various hours of the day. When nest B contained one egg, the bird was flushed at noon, returning in about 15 minutes with a bill full of grasses. It is believed that the females roosted there at this time, as nest II with four eggs was occupied at 10:15 p.m., and another with one egg at 5:30 a.m.
Incubation is performed by the female only, and begins normally\vith the laying of the last egg. At nests III and IX another egg was laid after incubation had apparently commenced. It may be significant that at both nests one egg hatched much later than the others, and this last young died before its first day was over.
During the first days, the female at nest I was absent for long periods, and on the morning of June 18, 2 days after the set had been completed, the eggs felt ominously cold. Twenty minutes later I found her incubating persistently, and thereafter she was seldom long absent. About two-thirds of the working day she spent on duty, a ratio she maintained throughout the period. While the percentage of time did not vary appreciably, it was noted that during late incubation the periods on and off the nest were of much shorter duration. These facts are indicated by the following figures, compiled from 3- and 2hour observations, respectively, at mid- and late incubation. Irrationally, the longest period away from the nest occurred during a brief cold shower.
6 days of ii days of incubalion incubation Time on nest 61.80 percent61.78 percent Time off nest 38.20 percent38.22 pAverage length of period on nest 21.6 minutes 7.6 minutes (max. 31) (maAverage length of period off nest 14.0 minutes 6.2 minutes (max. 33) (max. 12)
The incubating female crouches low in the nest with neck drawn in, wings laid compactly against the back, and tips of primaries crossed over the rump, only the bill and tail extending beyond the rim of the nest. This position is conspicuously different from the brooding posture, when she sits high over the nest, frequently panting from heat, neck outstretched, wings drooping over the rim of the nest.
At nest VII after one young had batched, the parent alternated these positions at frequent intervals.
During incubation the male continues to show a direct interest in home and mate, visiting the nest frequently. At nest III he appeared three times during 2 hours, and the pair flitted away together to feed. During the gray dusk which is the northern night, males of four nests were flushed from 20 to 60 feet from their nests. From the ground or the base of the bushes, they scudded low a few feet ahead of me, silently or with a low tsip. One sang a few bars of song and trailed off to silence in the middle of a note.
Young: Development of the young has been described in detail (Baumgartner, 1937). Summarizing the major points, hatching may occur at any time of the day, the sequence not dependent upon the order of laying, and the down dries in about 3 hours. The female subsequently swallows the egg shells.
Upon hatching the skin is yellowish with a pinkish undertone and is sparsely clad in tufts of fuscous down; bill and feet pinkish horn, lining of mouth orangish pink. The average length is 33.5 millimeters, weight 1.62 grams. Pin feathers begin to protrude at the age of 4 days on dorsal, ventral, and alar tracts, at 5 days on the caudal. At 6 days free feathers emerge beyond the sheaths of all tracts. At 8 days nestings are almost covered dorsally, the lower belly is bare, and the tail a stubby pincushion with feather tips barely showing.
At 9Y~ days, the age at which undisturbed young leave the nest, the back is fully covered, lower belly still slightly bare, wings twothirds grown, and tail still a stub; weight is 16.70 grams, a 930 percent gain over the bird’s weight on hatching, and equal to that of the average July adult.
Development of fledgling birds was traced in a brood confined to its nest area by a wire mesh cage, and through the recapture of banded young of known age. In both cases a loss of 1.5 grams was noted the first day out of the nest, then a gradual increase to 21 days with a total gain of 3 grams. Wings were then still slightly short and tails about two-thirds grown.
During August all birds were in heavy molt. In a series collected on August 19, measurements of the young were comparable to those of adults, and many were in almost complete adult plumage. Weights averaged slightly less than adults of the same sex.
IVei~,ht Weight Number Scx(in grams) Number Sex (in grams)
6 Male aduIt~.. 19. 7 14 Male immature 1&4 Female adult 17. 7 12 Female imma- 16. 8 ture 10 Average.~ iS. 7 26 Average 17. 5
Both parents shared the task of feeding their young, at first equally, later with the heavier load falling on the male. During the first few days the nestlings were fed only three or four times an hour; thereafter, an average of 16 feedings per hour was maintained, at intervals ranging from 1 to 10 minutes between visits.
During the early days the female brooded the young 74 percent of the time. Periods averaged about 8 minutes, occasionally as long as % hour, with intervals of only 3 to 8 minutes away from the nest. After a few days, feeding occupied a greater proportion of her time, and brooding periods were short and infrequent, rarely exceeding 3 minutes. At 6~4 days she was on 20 percent of the time during the very early morning, but not at all later in the day, though the female on nest IX was flushed from her nest on the 8th day. At night most of the birds remained with their young the full term of 93~ days.
At the age of 2 days the nestlings were sufficiently developed to stretch for food. From this time until fear was acquired, they responded with outstretched necks and gaping mouths to any motion. At 4 days the eyes were half open, and at the end of 5 were wide open, after which their responses were somewhat more discriminating.
They emitted no sound until the age of 5 days, and even at 8 days they usually simply raised their heads to accept what their parents brought.
Fear was acquired abruptly between 7~ and 8 days, correlating perfectly with the development of physical independence. In my daily rounds I had found them at first unalarmed by my presence and quiet during weighing. At nest III, where three banded young had hatched Y~ day before the other two, weighings were made just before these birds were 8 and 7~ days old, respectively. By keeping them in a hox they were retained long enough to weigh. As soon as they were returned to their nest, the three older birds scrambled off into the hushes, while the other two remained serenely their full term of 9~ days in the nest.
During the 9th day in the nest the young became quite active, shifting and stretching frequently, and greeting the parent bringing food by outstretched necks, fluttering wings, and noisy clamors. The undisturbed normal method of departure was observed at nest I. Arriving at the nest in the early afternoon of July 8, I found three of the brood already gone and the youngest, banded with a green thread at the time of hatching, sitting alone in the nest. The female bird came soon thereafter with food, which Green accepted quietly without moving to follow. His reaction was quite different when the male came. Immediately the young bird hopped past the proffered food and out onto the open tundra, peeping vigorously, sprawling and stumbling over the uneven ground 1 or 2 feet behind his father, who led him toward the nearby willow-birch thicket. About 20 feet from the nest he was fed, and thereafter he was given a morsel after every few feet, at intervals of 15 to 30 seconds. Occasionally Green wandered off the course and after considerable peeping by both parties, was finally found and fad. Thus it took them 20 minutes to traverse 100 feet of huminocky tundra. At the edge of the thicket the young bird was allowed to rest in the shelter of the hummock.
This performance was repeated almost identically when twice I brought him back to the nest. Green showed no desire to travel alone, nor to follow the female parent, whose instincts seemed to cling to the nest. Throughout my visit she chipped anxiously about it and fed her offspring several times while he was there. The third time that he was forcibly returned he concealed himself in the shrubbery directly behind the nest. The female came and did not see him, calmly swallowed the food herself and brooded for 2 minutes on the empty nest. Subsequently the male appeared and Green, now a true fledgling, hopped after him to the shelter of the thicket.
As has been indicated, young birds were quite unable to fly when they left the nest. The day following the departure from nest I described above, I found Green perched in a small tamarack about 15 inches from the ground in the same thicket to which I had followed him the previous afternoon. He was easily captured by hand, and upon his release he stumbled away over the uneven hummocks without trying to fly. Five (lays later a banded young from this same brood flushed 6 feet in front of me and flew laboriously some 30 to 40 feet. The development of caged young again corresponded with observations afield. Within the fenced corral droppings were first found on the branches a few inches above the ground level when the young were 15 days old, and they were thereafter conspicuously more active and quarrelsome. By the end of July young birds everywhere were flying about freely within the little thickets of their territory.
Fledglings over 2 weeks old, both at the cage and in the field, seemed to be fed as frequently as during their nest life. On July 26, when they were 22 days old, the male made three visits to the enclosure in ~ hour, although there was but one bird inside and three at large (the others had escaped). it was on this day that the young bird still in the corral gave the first indication of independent feeding. He was observed picking at the ground and was much attracted to a bread crust from my lunchbox.
Family groups farther afield were observed until August 15, but only occasionally was an adult secn gathering food for the young or feeding them.
On August 21, my last day in the field, the tundras were deluged with tree sparrows. Leaving the thickets where they had been reared, they were feeding now on the grass seeds of the open flats. The young were fully fledged, adults and immatures now indistinguishable, and the fall flocking had begun.
Plumages: The natal down has been described above, The juvenal plumage, worn by the fledgling until about 4 weeks of age, is as follows (Baumgartner, 1938): Pileum dull cinnamon brown, streaked by the black shafts of individual feathers, and more or less edged with light buff; nape similar, the black shafts indistinct or lacking, becoming grayer or buffer on sides of neck; feathers of back and scapulars broadly centered with black, edged with light buff and occasional touches of chestnut, especially the latter; rump and upper tail-coverts light buff, indistinctly st.reaked or mottled with black; sides of head and whole underparts light buff (shading to light smoke gray on throat and becoming a rich buff on sides), heavily streaked with soft black, especially on the breast, but not on lower abdomen and under tail coverts.
Except for the heavy streaking on the breast, the color pattern is essentially like that of the adult plumage, with the distinct pileum color terminated by a grayer nape, the broadly streaked back and rich buff of the sides. The same facial expression is produced by a light superciliary stripe, an irregular dark line under the eye, and a definite postocular streak. The dark pectoral spot, contrary to Beebe’s (1907) assertion, can be identified among the heavy streakings of the breast. A small patch of dull chestnut at the bend of the wing is streaked with black. The wings and tail are those of the first winter.
The first winter plumage is acquired by a partial post-juvenal molt in August which involves body plumage but apparently not the wings or tail, young and old becoming indistinguishable” (Dwight, 1900). This molt begins on the back and sides before the young are fully grown, about the first of August, and does not reach its culmination until after the middle of the month. The last juvenal feathers to disappear are those of the nape, the dark postocular stripe, and scattered streaky feathers of the throat and breast. None of the Churchill birds collected up to August 19 were in complete winter dress, and feet and bills were still pale.
When the species arrived in Ithaca in October, traces of the molt could still be discerned in 10 percent of those collected, of which 3 were young birds.
The nuptial plumage, for old and first year birds alike, is “acquired by wear, the buffy edging of the back becoming grayish and the chestnut everywhere slightly paler” (Dwight, 1900). A certain amount of this is due to fading, as Dwight intimates, but the chief change is by actual wearing away of the feather edges.
During the spring there is in addition a restricted molt of which Dwight (1900) says: “New feathers regularly grow on the chin in March, but apparently not in the other tracts and their appearance indicates, as in some other species, renewal rather than molt, for they are very few in number.” It was found, however, to be more extensive, for of 130 specimens examined, fresh sheaths were evident regularly on cheeks and chin, later on the throat, and other tracts were affected in the following proportions (in this count scattered feathers were not considered, but only definite areas of new quills): Number of Specimens Showing Tract Molt Crown (sides and back only) 5 Superciliary stripe 6 Lores 3 Nape 10 Dorsal tract 19 Scapulars 4 Rump 2 Sides Belly 6
Adult winter plumage is “acquired by a complete postnuptial molt, undistinguishab]e from first winter dress” (Dwight, 1900). At Churchill the first evidences, of this plumage were detected on July 81, 1933, and on Aug. 3,1934. Sequence was irregular, but in general contour feathers preceded the flight feathers, and the molt progressed from the anal region toward the head.
Although tree sparrows are described as “sexes identical,” a determined effort disclosed minor differences in plumage and measurements, a combination of which may distinguish up to 90 percent of birds at banding stations (Heydweiller, 1936). Using specimens which were later dissected to determine sex (by gonads) and age (by skull), a practical test with 129 birds gave the following score:
Male adult 36 birds, all judged correctly Male immature43 birds, S incorrect Female adult 19 birds, 7 incorrect Female immature 31 birds, all judged correctly Percentage of error 9.3 (due to overlap of characteristics of young males and adult females)
Criteria used in distinguishing sex and age are primarily lengths of wing and tail, width of crown, and amount of veiling of the crown. In general the largest birds are adult males, the smallest ones with distinct dark shafts on the crown feathers are first-year females. A wider crown usually indicates a male, a narrow crown a female. Male birds frequently show a loose flecking of buff over the whole crown, which in the female tends to be more heavily concentrated toward the center. The distinct dark shafts in the center crown feathers, though not invariable, are characteristic of first-year females. Operators of banding stations interested in sexing their tree sparrows should refer to the original article (Heydweiller, 1936) which gives measurements, a sketch of four typical crown patterns, and further details.
The western form, Spizella arborea ochracea, was recognized as a distinct subspecies by William Brewster in 1881. In describing it, he (Brewster 1882) remarks:
“The ground color of the back is decidedly paler than the ea~stern examples, bringing out the dark streaks in sharper contrast, which is heightened by the absence of their usual chestnut edging; ash of throat and sides of head is much fainter, in many places replaced by brownish fulvous; the underparts, especially the sides and abdomen, are more strongly ochraceous; and the broad ashy crown patch gives the head a very different appearance.”
The types of ochracea are now in the Harvard College Museum of Comparative Zoology: co-types from Fort Walla Walla, Washington, collected by Captain C. E. Bendire, male: Nov. 8,1881; female: Dec. 13, 1881.
Food: Beal (1897) found that a single tree sparrow eats about one-fourth ounce of seeds per day. Estimating conservatively that ten birds per square mile spend an average of 200 days in Iowa, then this species alone destroys 875 tons of weed seed annually. Tbis figure, he thought, could without exaggeration be multiplied by four. In the stomach of an individual bird he recorded 700 seeds of pigeon grass. A specimen I collected at Ithaca, N.Y., contained 982 seeds in the crop alone, with another 200 in a crushed mass in the stomach.
During its stay in the United States, the tree sparrow subsists almost entirely on weed and grass seed (Judd, 1901), with 98 percent seed food, about 2 percent animal matter, and an insignificant quantity of fruit. This species differs from associated fringillids in the large proportion of grass seed it eats, which makes up fully 50 percent of its vegetable diet. Panicums, pigeon grass, and allied grasses seem to be preferred, after which ragweed, lamb’s quarters, and the polygonums comprise another 40 percent, with the remaining 10 percent a variety of seeds, berries, buds, catkins, flowers, and waste grain.
Animal food found by Judd consisted chiefly of weevils and other beetles, ground beetles, rose beetles, wasp-like insects, ants, caterpillars, bugs, grasshoppers, and spiders. Knight (1908) adds flies, grubs and larvae. Hamilton (1933) found small stoneflies in the stomachs of four out of five tree sparrows collected in midwinter near a small stream near Ithaca, N.Y. He suggests that any warm day throughout the winter will bring thousands of transformed imagos to the vegetation surrounding such streams, and may account for the little flocks of birds that haunt such situations.
Knappen (1934), in summarizing the material in the Biological Survey files, states that in the 14 midwinter stomachs containing animal food, proportions varied from 1 to 90 percent of the total stomach content. Insects and spiders were found in adult form, as eggs, larvae, and pupae. Of a total of 550 stomachs on file, 38 contained animal food. During October animal matter comprised a third of the total. From November through March it ranged well below 10 percent. By April it had increased to almost a third again, and a single May specimen from North Dakota contained 100 percent animal food.
When they arrive on their northern breeding grounds, however, tree sparrows find conditions nearer winter than summer. Seeds continue to form the greater part of their diet until past the middle of June, when insects first become active (Baumgartner, 1937c). By mid-August the brief nesting season is over, and the proportion of seeds consumed again approaches the 98 percent maintained during the winter.
Young birds up to 3 or 4 weeks old are fed almost 100 percent animal matter, with a trace of Rosaceae leaves for their greens. When they begin to forage for themselves about the first of August, seeds and fruit are taken to a limited extent, their proportions gradually increasing. The last stomachs examined, on August 19, still showed a considerably larger proportion of animal food than did those of the adults of the same date.
A trace of gravel is first found in young birds of 3 days. At 5 days and thereafter gravel may compose from 10 to 15 percent of the stomach content. In August when they are feeding independently the gravel content increases to adult proportions, ranging from 15 to 70 percent, the majority about 35 percent.
In summarizing the summer food, it was noted that the outstanding vegetable item was the seeds of the genus Carex. These were found throughout the summer in all adults and juvenals that contained any vegetable matter, though they were not found in the nestlings. At least six species of Garex were distinguished in a single bird. In late July and August seeds of the crowberry (Empetrum nigrum), cranberry (Vaccinium vitus-idaea), and bulblets of the alpine knotweed (Polygonum viviparum), became an important part of the menu. Grass seeds, which form such a large proportion of the winter food were strikingly scant in the summer diet, being found in only one individual in late August. Besides seeds, plant matter was found in the form of soft bits of wood or chips, fibers, leaf fragments, galls, and unidentified debris.
The insects represent a large number of families and genera, among which the following groups are most conspicuous: Arachnida, Coleoptera (especially Donacia), Jchneumonidae, Lepidoptera, Nematinae, and Trichoptera. The adult stage predominated, but eggs, larvae, and pupae were also taken. A complete list of food species of the tree sparrow is included in Baumgartner (1937c).
In observations at the nest, the adult birds came repeatedly with bill overflowing with small green and brown caterpillars, as well as flies, mosquitoes, and other adult insects. At first the material is partially masticated, and the pulpy mass is lammed far down the young bird’s throat. Several birds may be fed at a single visit. Later great bilifulls of more or less entire insects are brought to the nest. But however great the quantity, it seems scarcely enough to satisfy one or two gaping mouths. The immensity of the old birds’ task was shown by a male collected in the field holding in its bill 41 adult insects, 4 larvae, and fragments of others, all captured presumably within 3 or 4 minutes.
Although during June and July in the North almost full daylight extends from 2:00 a.m. until 10:00 p.m., tree sparrows do not begin feeding until 3:15, and cease shortly before 9:00, a “working day” of 17.75 hours. From this it may be computed that, at an average of 16 feedings per hour, some 275 feedings are made daily, divided among the four or five young. It is interesting to compare with this the parental care given by that closely related species of more temperate latitudes, the chipping sparrow (Weed, 1.898). Averaging 12 visits per hour, divided among only 3 nestlings, each individual young of this species seems to be fed as many times per hour as do individual young of the tree sparrow. But here the working day begins at 5:00 a.m. and closes at 7:30 p.m. with only 14.5 hours in which to work. It is strongly suggested that these 3 extra hours a day make it possible for young tree sparrows to leave the nest at 9Y~ days, two days earlier than chipping sparrows at 11 ~ days. Thus the longer hours of daylight compensate in a measure for the briefness of the nesting season.
Economic importance: Because of the vast quantities of obnoxious weed seeds the tree sparrows consume during their winter sojourn in the States, much has been made of the economic value of this species. The Department of Agriculture estimates that the sparrow tribe: of which the tree sparrow is one of the most abundant species: saves the farmer $90 million a year. Judd (1901) describes the thoroughness with which they clean up a patch before moving on. On an area 18 inches square in a weedy ditch where they had been feeding, he found 1,130 half seeds, only 2 whole ones, and only 6 seeds left in the whole field, which, he says, was devoid of weeds the next year.
Since Judd’s time some doubt has been expressed of the value of the sparrow tribe. Certainly Judd overestimated the thoroughness of their gleanings, else they could not return year after year to the same areas, nor would they wander so freely over their little territories, only to cover the same ground another day. And certainly there is no scarcity of weeds in the country regardless of the great hordes of these birds. The reproductive capacities of the plants easily outdo the eating capacities of the sparrows, and there will probably always be enough weeds left to bother the farmer and propagate the species. Indeed, if there were no sparrows, the overcrowding of the plants themselves would soon establish a balance.
But if not actually beneficial, these birds are certainly harmless. They occasionally sample grain, but to no appreciable extent. The charge has been made that they distribute rather than destroy the seeds, hut this accusation was refuted by Judd’s study. He found that in the thousands of stomachs containing ragweed, there was never an unbroken seed. The thoroughness of avian digestion prevents the evacuation of anything but a most insignificant portion of the food ingested.
In the summer the tree sparrow is of no economic significance, as it nests beyond the reaches of civilization. But whether or not we can evaluate the species in cold dollars and cents, it will always be welcome as a gentle, cheerful little creature in our winter fields and gardens.
Behavior: Since the earliest days of nature lore in America, writers of the winter fields have thrilled to the cheerful warble of these hardy little visitors from the North. I shafl never forget my first flock of tree sparrows, feeding companionably at the weedy border of the marsh, hanging on the weed tops like animated Christmas tree ornaments, dropping lightly to the ground and etching their delicate tracery of claw prints in the snow. The air was mellow with their soft warbles, and to me they have always said “Marguerite, Marguerite.” Erect, dignified, they are at some times more vivacious than others. On stormy days in midwinter they may huddle quietly in the lee of a wall, while in the springtime, or indeed any sunny day, we may find them playing a gay game of tag through the shrubbery.
Keynotes of the t.ree sparrow’s character are gentleness and unsuspiciousness. Easily baited in large numbers, quiet in the hand, and a frequent repeater, this species has long been a favorite at banding stations. Both at feeding stations and on the nest.ing territory they are amiable and unaggressive, going their quiet way without the nervousness and fussiness of so many species.
When flushed, tree sparrows swoop up into the nearest bush or tree and perch quietly from 3 to 10 or 12 feet from the ground. If the intruder stands quietly, they will soon forget him and return to their endless banquet. During the migration they tend to be more shy and secretive, flushing sooner and flying greater distances. In summer the incubating female flushes usually at 4 to 6 feet, and reappears a moment later, chipping with gentle anxiety from the top of a scrub birch a few feet away. The male, less nervous, less vociferous in his protests at an intruder, is more direct and presumptive about the nest. Even in feeding the young, when duties of the sexes are identical, he can easily be distinguished by his perkier air, erect crown, and more direct manner of approaching the nest, while the female fusses and chips and finally creeps through the grasses to her young.
The tree sparrow’s flight is unhurried, graceful, and slightly undulating. In a normal day’s foraging, the birds fly no great distances, drifting through bedgerows or marshes, passing and repassing one another in their irregular advances. When traversing pronounced distances across bare areas, the flock rises with a cirding, swirling movement to about 100 feet and sails off in irregular formation to points unknown.
When food is abundant or winds are biting, they may perch quietly for 20 to 30 minutes at a time with neck drawn in, tips of the primaries slightly crossed over the rump, and contour feathers fluffed to twice their nornial size, almost covering wings and legs. For such extended rests a small limb in denser shrubbery is usually chosen, from three to ten feet from the ground. For short stops any twig will do, usually near the ground, but not infrequently in the higher tree tops.
As their winter diet consists principally of weed seeds, tree sparrows feed largely on the ground, scratching among the dry grasses or hopping up at bent-over weed-heads. They are less inclined to dig than song sparrows, and when food is plentiful will pass lightly over the more obvious patches. Often one balances on a weed top, swaying in the wind and scattering the seeds on the snow below, where they are promptly appropriated by his companions. That one bird deliberately performs this function for the others, as has been sentimentally inferred, is unlikely. On the contrary, it has been observed that the foraging birds rarely encroach within six inches of one another, and maintain small and definite feeding territories for themselves.
In the early summer at Churchill they were seen, usually in pairs, feeding about the grassy edges of little pools and, not infrequently, picking the fresh buds and catkins off the stunted willows and birches. These they obtained either by perching on the branch or by hopping up from the ground, sometimes to a height of several inches. Later, when the young had hatched and the diet had changed to chiefly animal food, the parents were seen gleaning caterpillars and small insects among the leaves and branches of the nearby thickets. Occasionally one darted into the air for a mosquito or moth, but these forays were short, and if the prey escaped it was not ardently pursued.
Water is as essential as food, and in winter when there are no open pools, I have frequently watched tree sparrows about my station swallowing snow, either from the ground or from the crotches of trees. They enjoy bathing even in the coldest weather, nor are they content merely to wade, but hop to their middles and splash and duck and flutter like diminutive ducklings.
While tree sparrows are gregarious at night as well as by day, they seem to prefer a solitary roost, for tramping through their haunts frequently after dark I flushed only scattered individuals. I believe they prefer cattail marshes when these are available. Here they creep into little cavities on or near the ground, protected from storms, wind, and enemies. Weedy fields, corn shocks, and haystacks are also popular. Dense growths of young white pine offer shelter not unlike their northern home. A flock of a hundred or more I watched near Ithaca, N.Y. usually assembled in the late afternoon in a weedy corn patch, and at dusk drifted up the valley to a stand of young pines over a half mile away. Straggling in loose formation, yet with some definiteness of purpose, they spread out over the whole area so that only single individuals could subsequently be flushed.
Voice: Across the barrens of the North, the little song of the tree sparrow rings as clear and sweet and simple as one who loves its gentle ways could desire. Though quite different in pattern, its song resembles that of the field sparrow in pitch and intensity, and it has that silver, bell-like quality that is at the same time both plaintive and serene. There is also a decided resemblance in accent and rhythm to the final notes of the chestnut-sided warbler. In length its song ranges from 3)~ to 4 seconds, with an occasional rollicking variation of only 1~ seconds.
Some of the variations in my field note book are: tsee tsee-ah, tsi tsit su; tsee tsee: ah, tsit sut sut; t.see tsce-ah, tsi tseedle-cedle: ali; tsee tsweee ~Si, t~i tS?mt 8U trtrtrtrtr (end a clicking trill) (these are variations at a single nest); tsi tsi t8weeeee, t8wce t.swce tswee tswit list t: sut, and a brief warble-like tse-weet, tse: weet, tse-wi-tse-tse-tswegt.
During Juno the nnUes sing continuously, one song following another in rapid succession. On one occasion I counted four in a minute, followed by a brief interval whilc tho bird flew to another perch. There is no siesta period in this cool climate. Night singing, in spite of the half dark twilight of the midnight sun, is infrequent. On several occasions when I watched nests all night, singing decreased conspicuously after 9:15 and the last song was heard at 10:15 p.m., although other species were still active. Although there was full daylight at 2:15 am, the first tree sparrow song was not recorded until 2:45.
In mid-July singing becomes less frequent as males take over their share of feeding the young. During August the song can be heard occasionally on warm still days.
During fall migration the song is heard not infrequently during November, a thin, wiry note that rarely reaches the fullness of the true breeding song. February is the beginning of the spring tune-up, and by mid-March the hedgerows ring with the true summer song. With t.he passing of most of the males, singing becomes less frequent after the middle of April.
That melodious, warbling twitter of a winter flock which Thoreau (1910) so aptly describes as the “tinkle of icicles” cannot, according to Saunders (1929h) be considered true song, as it is not a courtship performance but merely the contented utterance of a well-fed flock. It can be heard at any time and in any weather throughout the winter a cheerful, tri-syilablic warble, teedle eet, teedle eet.
Call notes include the soft trip on winter feeding grounds, a somewhat sharper alarm note heard both winter and summer, an imperious little breeding call of the male clielc-chelc-ckek, chek, chek, the female’s response, a soft whey-whey-whey-whey-whey, and whispered ts-ts-ts-ts-ts. Sometimes when the male came to feed the young, the brooding female uttered rapidly and with rising inflection, a series of consonants which I recorded in by field notebook as “pppppppt,” or a low ticking note. Sometimes both parents came to the nest simultaneously and stood fondly over their family, uttering low cooing notes and touching bills tenderly.
Development of voice in the young was traced from the first soft tseeeeeeee at 5 days (Baumgartner, 1938a) to the noisy food call at 9Y~ days as the young left the nest, when both adult and nestling uttered a raucous aah aah aah aah.
Enemies: Man, the archenemy of many birds, affects the tree sparrows very little. Too small for game, beneficial in habits, inconspicuous in manner, they are seldom noticed in winter, and in summer retreat mainly beyond the reaches of civilization. Predatory animals, such as the bird hawks, owls, shrikes, weasels, and the domesticï cat account for a few in winter. In summer they are beyond the domain of snakes, cats, most crows, jays, and squirrels, and such corn petitors as English sparrows, starlings, and cowbirds. I found nest mortality and infertile eggs to be 21.3 percent.
Parasites, both external and internal, were found to be relatively few (Baumgartner, 1937a). Of some 600 specimens carefully in spected, many were entirely free and none harbored more than a scattering. Of 26 nests observed at Churchill, none showed evidence of infestation.
The most serious enemies of this species are the elements: snows that cover the food supply, and storms and sudden cold spells, particu larly during the long migration. While the birds can withstand tem peratures to : 28~ C. (West, 1960), they cannot survive without food.
A late traveler in the fall, an early one in spring, they run the gantlet of the equinoctial storms at both seasons. W. E. Saunders (1907) counted 358 dead tree sparrows in 3 hours after an October storm on Lake Huron.
Of the thousands upon thousands of this species that have been handled at banding stations, only a small porportion survive to their 4th year. A handful have returned a 5th and 6th year, and the oldest birds on record are a return-8 taken by B. S. Bowdish of Demarest, N.J. (in litt.) and another by Paul Nighswonger (1959) at Alva, Okla. Assuming that these birds were banded in their first winter, they would have been between 8~ and 9 years old on last appearance.
Survival ratio of tree sparrows has been computed by Baumgartner (1937a) based on nest mortality, proportion of first year and adult specimens collected through the season, and banding returns, both personal and those recorded in the literature.
Calculating the fate of a potential 750 eggs from 150 nests:
78.7 percent survive the nestling period =590 birds 50 percent of these 590 survive the first migration and winter =295 birds 40 percent of these 295 make winter Rl = 119 birds 43 percent of these 119 survive to R2 : 51 birds 34 percent of these 51 survive to R3 = 18 birds 28 percent of these 18 survive to R4 : 5 birds 16 percent of these 5 survive to R5 : 1 or 2 birds By the sixth summer only one old veteran remains, who may live one or two more years.
Fall and Winter: Beginning the southward trek in September, the hordes of tree sparrows reach southern Ontario about the first of October (Taverner and Sutton 1934) and by the end of the month are flooding the States. They normally reach Oklahoma, near the southern part of their range, by early November and remain until March.
Traveling, apparently, in family and neighborhood groups, the flocks upon arrival at Ithaca, N.Y., were composed of almost equal proportions of males and females, with a 16 percent dominance of males (Baumgartner 1937a). First-year birds (with skulls incompletely ossified) exceeded old birds 3 to 2 between mid-October and mid-December. During the non-migratory period between January and March, the proportion dropped to almost equal numbers of young and old birds, indicating a higher mortality of first-year birds. Females, traveling farther south, dropped to a 3 to I proportion until near the end of March.
The extent of winter territory and flocking habits has been the subject of several intensive studies, foflowing different techniques and with a variety of conclusions. At Ithaca, N.Y., in 1933, 1934, and 1935, I marked individuals at four stations by gluing different colored feathers to their tails (Baumgartner, 1938b). By following these marked individuals and by retrapping at other stations, I found that the normal feeding range in winter was 500 to 1,700 feet in diameter, average 1,000 feet. Extensions from 3,000 to 0,800 feet, recorded for 40 percent of the birds, were associated with fall settling, spring excitement, or heavy snowfalls. A few individuals appeared to be regular wanderers.
Winter flocks appeared to be loosely defined units of varying numbers and individuals, with a fairly definite flock territory, from which, however, individuals strayed at will, flocks inhabiting open country ranged more widely than those in denser cover such as marshes. In severe weather, flocks split into smaller groups and wandered more widely.
The histories of individually marked birds suggest several explanations for low return ratios at banding stations. Besides migrants, which are unlikely to make the same stop-overs a 2nd year, this study recorded several of each of the following types:
(1) Trap-shy individuals that were caught only once or twice a season, though observed regularly in the vicinity.
(2) Wanderers that covered so large an area they missed their original station another season.
(3) Regular residents of another area that meandered only once into the trap where banded.
(4) Emigrants that shifted from year to year.
A project at Amherst, Mass., during the winter 1957: 58 (Sargent, 1959) used colored bands at five stations. Sargent likewise considered flock structure unstable, with no fixed associations within a flock, or regular times of day for visitations to the traps. lie also attributed wandering to winter storms, with consequent influxes of new birds to a station. Fifteen of his birds wandered over $ mile, seven wandered over 2 miles and one to a point 7% miles away. He suggests that the social organization, described as a straight-line pecking order modified by reverse peeks (Sabine, 1949), may be partially responsible for wandering, as over-crowding at a feeding station might induce social intolerance and force away individuals near the bottom of the social hierarchy.
Helms and Drury (1960), working at South Lincoln, Mass., colorbanded 477 tree sparrows, with almost 2,000 handlings to gather data on measurements, weights, and fat deposition. Field observations were made on a fairly stable winter population of about 50 birds to determine size, behavior, composition, and changes of winter flocks. They describe foraging groups as loosely integrated groups, usually four to eight birds, traveling and feeding together for most of the winter. The reason these birds do not coincide regularly in the traps, they believe, is because the foraging group remains only 10 to 20 niimites in a feeding area, and thus the likelihood of recapture together is slight. During a normal winter day, foraging individuals spend a maximum of 6 to 10 minutes in feeding activities, followed by a 12- to 20-minute period of either perching, preening, bathing or social activity that may carry the group to another foraging area.
Tree sparrows appear to have a strong homing instinct. Not only do they return year after year to banding stations; experiments in transporting birds varying distances indicate that they return with astonishing rapidity (Ileydweiller, 1935). A bird transported 5 miles was back the following morning. An individual taken 10 miles covered the intervening fields in 16 days. Of 14 birds carried from 17 to 100 miles, 3 returned the following winter. A fourth was collected the following year from a flock 3 miles from the station.
Sargent (1959) transferred six color-banded regular winter residents of one of his stations 7% miles to another. Three were retrapped at the original station from 2 to 18 days later. Another traveled an additional 3% miles in the opposite direction from its original area, and was observed twice at this point. On February 9, he similarly transferred 20 birds new to the station following a snowfall. Some remained at the new station for about a nionth. None returned to the original station, for which, presumably, they had no home attachment.
DISTRIBUTION
Eastern Tree Sparrow (S. a. arborea)
Range: Mackenzie, Keewatin, northern Ontario and Labrador south to Oklahoma, rrennessee and Virginia.
Breeding Range: The eastern tree sparrow breeds in central and eastern Mackenzie (mouth of Coppermine River, Fort Rae, Hanbury River), northern Saskatchewan (Fort Fond du Lac, Reindeer Lake), central western Keewatin (bare Lake), northeastern Manitoba (Churchill, Cape Tatnam), northern Ontario (Fort Severn, Little Cape), northern and central eastern Quebec (Lake Minto, Fort Chimo, Paint hills, Old Romaine, Bradore), and Labrador (Okak, Battle Harbor).
Winter Range: Winters from central Minnesota (Sherburne and Isanti counties), Wisconsin, northern Michigan (Schooleraft and Luce counties), central Ontario (North Bay), southwestern Quebec (Montreal), Maine, New Brunswick, Prince Edward Island, and Nova Scotia south to Oklahoma, Arkansas (Winslow), Tennessee (Memphis, Nashville), western North Carolina (Asheville, WinstonSalem), and Virginia (Blacksburg, Quantico); casually south to southern Nevada (Lake Mead), southeastern Texas (Hardin), northwestern Mississippi (Rosedale), central South Carolina (Summerton), and Bermuda.
Migration: The data deal with the species as a whole. Early dates of spring arrival are: Maine: Bangor, March 20. Quebec: Montreal area, March 17 (median of 8 years, March 22). Illinois: Chicago, February 26 (average of 16 years, March 15). Ontario: Ottawa, average of 18 years, April 12. North Dakota: Cass County, February 23 (average, March 10).
Late dates of spring departure are: Alabama: Gadsden, March 16. North Carolina: Cherokee County, March 9. Virginia: Lexington, April 9. District of Columbia: April 24 (average of 29 years, March 27). Maryland: Prince Georges County, April 14 (median of 6 years, March 28); Allegany County, April 12. Pennsylvania: State College, May 22. New Jersey: Troy Meadows, May 12 and May 5. New York: Ontario County, May 11; Westchester County, May 8. Connecticut: South Windsor, May 3; New Haven, April 28. Massachusetts: Nantucket, May 29; Martha’s Vineyard, May 3 (median of 6 years, April 10). New Hampshire: New Hampton, May 12 (median of 21 years, April 25); Concord, April 30. Maine: Bangor, May 16. Quebec: Montreal area, May 9 (median of 14 years, April 30). New Brunswick: May 28. Nova Scotia: Wolfville, April 30. Arkansas: Fayetteville, April 3. Tennessee: Athens, April 22 (average of 5 years, April 8); Nashville, March 27. Kentucky: Lexington, March 30. Missouri: St. Louis, April 28 (median of 15 years, April 20). Illinois: Chicago, May 10 (average of 16 years, April 26); Urbana, April 25 (median of 19 years, April 8). Indiana: Wayne County, April 14 (median of 11 years, April 6). Ohio: Buckeye Lake, May 7 (median of 40 years for central Ohio, April 20). Michigan: Detroit area, May 13 (mean of 10 years, May 7); Battle Creek, May 2 (average of 30 years, April 17). Iowa: Sioux City, April 21 (average of 32 years, April 1). Minnesota: Minneapolis-St. Paul, April 25 (mean of 6 years, April 8). Kansas: northeastern Kansas, April 14 (median of 14 years, April 4). Nebraska: Red Cloud, April 20. South Dakota: Sioux Falls, May 7 (average of 8 years, April 23). North Dakota: Cass County, May 5 (average, April 29). Wyoming: Laramie, April 14 (average of 9 years, April 6). Idaho: Lewiston, April 10 (median of 11 years, April 1).
Early dates of fall arrival are: Idaho: Lewiston, October 16 (median of 11 years, November 1). Wyoming: Cheyenne, September 28 (average of 11 years, October 17). New Mexico: Rio Grande Valley, November 6. Manitoba: Treesbank, September 9 (average of 21 years, September 26). North Dakota: Jamestown, October 1; Cass County, October 1 (average, October 6). South Dakota: Sioux Falls, October 4 (average of 7 years, October 19). Nebraska: Red Cloud, October 11. Kansas: northeastern Kansas, October 7 (median of 16 years, October 21). Texas: Fort Worth, November 14. Minnesota: Minneapolis-St. Paul, October 5 (mean of 12 years, October 17). Iowa: Sioux City, October 8 (average of 32 years, October 15). .N’Iichigan: Detroit area, September 25 (mean of 10 years, October 7); Battle Creek, October 15 (average of 29 years, October 27). Ohio: central Ohio, September 27 (median of 40 years, October 20). Indiana: Wayne County, October 8 (median of 10 years, November 19). Illinois: Chicago, October 1 (average of 16 years, October 10). Missouri: St. Louis, October 10 (median of 15 years, October 25). Kentucky: Bowling Green, November 7. Tennessee: Clarksville, November 21. Arkansas: Fayetteville, October 18. Nova Scotia: Margaretsville, October 23 (average of 11 years, October 27). New Brunswick: St. Andrews, September 16; Scotch Lake, September 23. Quebec: Montreal area, September 27 (median of 14 years, October 13). Maine: Lake Umbagog, October 9. New Hampshire: New Hampton, October 15 (median of 21 years, October 21); Concord, October 21. Massachusetts: Martha’s Vineyard, October 5 (median of 5 years, October 19). Connecticut: East Windsor Hill, September 30; Farmington, October 1. New York: New York City, September 16; Ontario County, September 17. New Jersey: Island Beach, October 4. Pennsylvania: State College, October 1; Renovo, October 16. Maryland: Allegany County, October 18; Laurel, November 6 (median of 5 years, November 7). District of Columbia: October 3 (average of 16 years, November 10). Virginia: Shenandoah National Park, October 31. North Carolina: Arden, November 25.
Late dates of fall departure are: Manitoba: Treesbank, November 15 (average of 20 years, November 6). North Dakota: Cass County, November 24 (average, November 12). Illinois: Chicago, December 7 (average of 16 years, November 17). Newfoundland: St. Anthony, October 9. Prince Edward Island: Murray Harbour October 26. Quebec: Montreal area, November 19. Maine: Lake Umbagog, November 9.
Egg dates: Alaska: 128 records, May 27 to June 29; 68 records, June 4 to June 16.
Labrador: 16 records, June 10 to July 11; 11 records, June 20 to June 29.
Manitoba: 14 records, June 12 to June 26. Ontario: 1 record, July 1.
Western Tree Sparrow (S. a. ochracea)
Range: Northern Alaska, Yukon, and northwestern Mackenzie south to northeastern California, Arizona, New Mexico, and central Texas.
Breeding Range: The western tree sparrow breeds from northern Alaska (Kobuk and Colvile deltas), northern Yukon (Firth River), and northwestern Mackenzie (Mackenzie Delta, lower Anderson River) south to southwestern and central Alaska (Naknek, Wrangell Mountains), northwestern British Columbia (Atlin), southeastern Yukon (Sheldon Lake), and central western Mackenzie (Fort Franklin).
Winter Range: Winters from southern British Columbia (Vancouver, Okanagan Landing), southwestern Saskatchewan (Cypress Hills), South Dakota (Harding and Hutchinson counties), and northcentral Iowa (Sioux City) south to northeastern California (Fort Creek, Wendell), central western Nevada (Carson City), northern and central eastern Arizona (San Francisco Mountains, San Carlos), central New Mexico (San Antonio), and central Texas (San Angelo, Giddings); casually north to Alaska (Fairbanks, Wrangell) and south to southern California (Riverside, Death Valley).
Casual records: Casual north to Banks Island.
Egg dates: Mackenzie: 29 records, June 6 to July 4; 14 records, June 14 to June 21.